




































Science 重磅!科学家发现氨氧化古菌AOA在无氧条件下产生氧气和氮气
发布日期:2022-01-15 浏览次数:274
众所周知,氨氧化古菌(AOA)是一类广泛存在于海洋中的微生物,负责将氨氧化生成亚硝酸盐,在地球氮元素循环具有重要作用。然而,在氧气缺乏的环境中,如海洋沉积物、黑海中,AOA的丰度也很高,这意味着AOA可能存在某种适应机制得以在无氧条件下生存繁衍。来自南丹麦大学的Beate Kraft研究团队就这一问题展开探索,并将研究成果整理发表于Science:”Oxygen and nitrogen production by an ammonia-oxidizing archaeon”。
一、N. maritimus在无氧条件下生成氧气
Beate Kraft博士首先在有氧条件下对N. maritimus进行无菌培养,然后通入氩气使得培养液中氧水平降低到5µM以下,残余的氧气被N. maritimus氨氧化作用所消耗。研究者通过痕量发光氧传感器探测孵育体系中的氧气浓度,令人惊奇的发现当体系中氧气被完全消耗到检测线1nm以下后,氧浓度反而缓慢地增加。研究者又设计了一系列重复实验,通过给N. maritimus孵育体系中补充氧饱和水,会发现氧气先逐渐被消耗,氧浓度降低直至检测线下,然后氧浓度反而增加,在不再添加氧的情况下能达到100~200nM。当过滤除去生物质或加入氯化汞杀死细胞后,则无法观测到氧气水平积累的现象,同时研究者也进一步排除了其他潜在的干扰因素。另外,孵育体系中少量的NO被研究者运用微电极探测到,且会干扰O2的光极测量,但不会对O2微电极探测造成干扰。研究者运用光极探测O2、微电极探测NO就可以纠正O2水平,得到其真实浓度,但这个校正值其实相当微小(0-17%)。研究还证明氨氧化和亚硝酸盐转化的其他中间体或副产物对氧光极探测氧浓度不存在干扰。
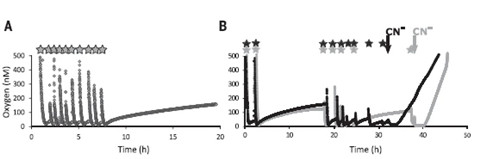
图1N. maritimus在厌氧条件下生成氧气
二、N. maritimus中存在氧气生成与耗的两个平衡反应
研究者大胆提出假设在N. maritimus中存在两个由氨氧化反应偶联的氧气生成和氧气消耗的平衡反应过程,氧气的净积累是两个反应过程随着时间平衡的结果。为了验证这个假设,研究者在培养液中添加抑制氨氧化作用的氰化物(0.5mM),结果发现当添加氰化物后,氧浓度出现初始短暂的停滞期,然后以相对于添加前5倍的速率平稳增加,同时添加氰化物后,没有探测到NO的生成。据此,研究者提出的假设得以证实,N. maritimus中确实存在生成氧和消耗氧的两种生化反应,没有氰化物的情况下,一部分胞内生成的氧气被用于其他反应而没有释放到环境中。研究者运用15N示踪技术,设计两组平行实验:15NH4+50µM分别与5µM14NO2-(孵育体系Ⅰ)和 25µM14NO2-(孵育体系Ⅱ)发现随着胞内氧气生成,氨不断被氧化生成亚硝酸盐。进一步研究者测量两个孵育体系的氨氧化反应速率为46nM/h、36nM/h,所需氧生成速率为69nM/h、60nM/h,由此研究团队得出化学计量方程式:(NH3+ 1.5O2 NO2− +H2O + 1H+)。研究者测量发现两个体系中,氧积累平均速率都只有1.2nM/h,由此,证实N. maritimus胞内生成的氧很快就被氨氧化反应。
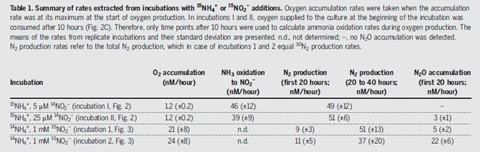
图2N. maritimus在氧气生成过程还生成亚硝酸盐和N2
三、N. maritimus在氧气生成过程还生成亚硝酸盐和N2
研究者进一步对胞内氧气代谢途径展开了探究,通过抽丝剥茧最终发现N. maritimus在有氧条件下将氮转化生成NO,NO通过歧化作用可能生成氧气。甲烷氧化菌NC10中存在NO歧化生成氧,然后再氧化甲烷,由于氧的生成与消耗是紧密偶联的,目前尚未可知甲烷氧化NC10细菌是否能释放多余的氧气到环境中。研究者用NO微电极发现NO和氧气产生是偶联产生的,但偶联关系在某些条件下并不是很强,当添加氰化物,则完全解偶联,但是如果添加NO清除剂PTIO,氧气产生也中止。综上,研究者证实NO是氧气产生过程中的重要中间体。
四、在N. maritimus中氧气、氮气生成与N2O积累同步发生
研究者根据两组孵化实验的探测结果,推论得到在N. maritimus中NO是N2和O2产生的中间体,NO首先通过亚硝酸盐产生,然后歧化生成N2和O2,在15N标记亚硝酸盐孵育体系1和2中,前20小时氧气的积累量超过氮气,表明在这个阶段氧气和氮气生成是不同步的。由前面的研究结果已知,O2积累的净速率是低于总生成速率的,因此N2和O2的生成速率一定存在一个不平衡。如果真是这样,那么O2积累的速率应该不会超过N2产生的速率,但是通过测量得到计量数据表明在NO 、N2和O2之间可能还存在一个反应中间体N2O,关系式为:2NON2O+0.5 O2。事实上,研究者在15N标记的亚硝酸盐实验中观测到46N2O 积累早于30N2生成,在热力学上NO歧化反应生成N2O和O2也是有利的。
单纯就NO作为中间体歧化生成N2、O2而言,NC10和N. maritimus是类似的,然而研究结果表明N. maritimus还存在一个额外的中间体N2O,并与氧气瞬时积累有关。综上,研究者给出N. maritimus氧气生成的可能的代谢途径模式图,见图三。尽管在N. maritimus中,氨氧化亚硝酸盐的氧气可以自给自足,但是反应所需的电子并不完全依赖于铵根离子,有氧氨氧化过程中产生的胞内或胞外的有机质,或者是水体及其分解产生的有机质可能也作为反应的电子供体。
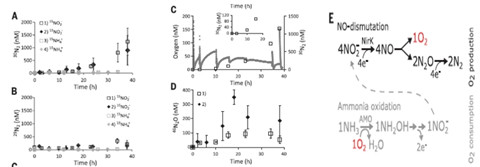
图3 在N. maritimus中氧气、氮气生成与N2O积累同步发生
五、总结与展望
本研究成果拓宽了人们对AOA在厌氧环境中的功能认知,Beate Kraft博士说N. maritimus在氧气消耗殆尽后,还能产生N2和O2,揭示了环境中N2生成的一类重要途径。当前的15N示踪技术运用于测量海洋厌氧环境中N循环速率可能忽视了这种途径,并将其 N2和 N2O 生成归结为典型反硝化和/或厌氧氨氧化。鉴于全球海洋中AOA的高丰度及其在N循环中的关键作用,广泛理解AOA的功能可以加深人们对乏氧环境中的微生物生态学和生物地球化学循环的认知。
转载自腾讯网
众所周知,氨氧化古菌(AOA)是一类广泛存在于海洋中的微生物,负责将氨氧化生成亚硝酸盐,在地球氮元素循环具有重要作用。然而,在氧气缺乏的环境中,如海洋沉积物、黑海中,AOA的丰度也很高,这意味着AOA可能存在某种适应机制得以在无氧条件下生存繁衍。来自南丹麦大学的Beate Kraft研究团队就这一问题展开探索,并将研究成果整理发表于Science:”Oxygen and nitrogen production by an ammonia-oxidizing archaeon”。
一、N. maritimus在无氧条件下生成氧气
Beate Kraft博士首先在有氧条件下对N. maritimus进行无菌培养,然后通入氩气使得培养液中氧水平降低到5µM以下,残余的氧气被N. maritimus氨氧化作用所消耗。研究者通过痕量发光氧传感器探测孵育体系中的氧气浓度,令人惊奇的发现当体系中氧气被完全消耗到检测线1nm以下后,氧浓度反而缓慢地增加。研究者又设计了一系列重复实验,通过给N. maritimus孵育体系中补充氧饱和水,会发现氧气先逐渐被消耗,氧浓度降低直至检测线下,然后氧浓度反而增加,在不再添加氧的情况下能达到100~200nM。当过滤除去生物质或加入氯化汞杀死细胞后,则无法观测到氧气水平积累的现象,同时研究者也进一步排除了其他潜在的干扰因素。另外,孵育体系中少量的NO被研究者运用微电极探测到,且会干扰O2的光极测量,但不会对O2微电极探测造成干扰。研究者运用光极探测O2、微电极探测NO就可以纠正O2水平,得到其真实浓度,但这个校正值其实相当微小(0-17%)。研究还证明氨氧化和亚硝酸盐转化的其他中间体或副产物对氧光极探测氧浓度不存在干扰。
图1N. maritimus在厌氧条件下生成氧气
二、N. maritimus中存在氧气生成与耗的两个平衡反应
研究者大胆提出假设在N. maritimus中存在两个由氨氧化反应偶联的氧气生成和氧气消耗的平衡反应过程,氧气的净积累是两个反应过程随着时间平衡的结果。为了验证这个假设,研究者在培养液中添加抑制氨氧化作用的氰化物(0.5mM),结果发现当添加氰化物后,氧浓度出现初始短暂的停滞期,然后以相对于添加前5倍的速率平稳增加,同时添加氰化物后,没有探测到NO的生成。据此,研究者提出的假设得以证实,N. maritimus中确实存在生成氧和消耗氧的两种生化反应,没有氰化物的情况下,一部分胞内生成的氧气被用于其他反应而没有释放到环境中。研究者运用15N示踪技术,设计两组平行实验:15NH4+50µM分别与5µM14NO2-(孵育体系Ⅰ)和 25µM14NO2-(孵育体系Ⅱ)发现随着胞内氧气生成,氨不断被氧化生成亚硝酸盐。进一步研究者测量两个孵育体系的氨氧化反应速率为46nM/h、36nM/h,所需氧生成速率为69nM/h、60nM/h,由此研究团队得出化学计量方程式:(NH3+ 1.5O2 NO2− +H2O + 1H+)。研究者测量发现两个体系中,氧积累平均速率都只有1.2nM/h,由此,证实N. maritimus胞内生成的氧很快就被氨氧化反应。
图2N. maritimus在氧气生成过程还生成亚硝酸盐和N2
三、N. maritimus在氧气生成过程还生成亚硝酸盐和N2
研究者进一步对胞内氧气代谢途径展开了探究,通过抽丝剥茧最终发现N. maritimus在有氧条件下将氮转化生成NO,NO通过歧化作用可能生成氧气。甲烷氧化菌NC10中存在NO歧化生成氧,然后再氧化甲烷,由于氧的生成与消耗是紧密偶联的,目前尚未可知甲烷氧化NC10细菌是否能释放多余的氧气到环境中。研究者用NO微电极发现NO和氧气产生是偶联产生的,但偶联关系在某些条件下并不是很强,当添加氰化物,则完全解偶联,但是如果添加NO清除剂PTIO,氧气产生也中止。综上,研究者证实NO是氧气产生过程中的重要中间体。
四、在N. maritimus中氧气、氮气生成与N2O积累同步发生
研究者根据两组孵化实验的探测结果,推论得到在N. maritimus中NO是N2和O2产生的中间体,NO首先通过亚硝酸盐产生,然后歧化生成N2和O2,在15N标记亚硝酸盐孵育体系1和2中,前20小时氧气的积累量超过氮气,表明在这个阶段氧气和氮气生成是不同步的。由前面的研究结果已知,O2积累的净速率是低于总生成速率的,因此N2和O2的生成速率一定存在一个不平衡。如果真是这样,那么O2积累的速率应该不会超过N2产生的速率,但是通过测量得到计量数据表明在NO 、N2和O2之间可能还存在一个反应中间体N2O,关系式为:2NON2O+0.5 O2。事实上,研究者在15N标记的亚硝酸盐实验中观测到46N2O 积累早于30N2生成,在热力学上NO歧化反应生成N2O和O2也是有利的。
单纯就NO作为中间体歧化生成N2、O2而言,NC10和N. maritimus是类似的,然而研究结果表明N. maritimus还存在一个额外的中间体N2O,并与氧气瞬时积累有关。综上,研究者给出N. maritimus氧气生成的可能的代谢途径模式图,见图三。尽管在N. maritimus中,氨氧化亚硝酸盐的氧气可以自给自足,但是反应所需的电子并不完全依赖于铵根离子,有氧氨氧化过程中产生的胞内或胞外的有机质,或者是水体及其分解产生的有机质可能也作为反应的电子供体。
图3 在N. maritimus中氧气、氮气生成与N2O积累同步发生
五、总结与展望
本研究成果拓宽了人们对AOA在厌氧环境中的功能认知,Beate Kraft博士说N. maritimus在氧气消耗殆尽后,还能产生N2和O2,揭示了环境中N2生成的一类重要途径。当前的15N示踪技术运用于测量海洋厌氧环境中N循环速率可能忽视了这种途径,并将其 N2和 N2O 生成归结为典型反硝化和/或厌氧氨氧化。鉴于全球海洋中AOA的高丰度及其在N循环中的关键作用,广泛理解AOA的功能可以加深人们对乏氧环境中的微生物生态学和生物地球化学循环的认知。
转载自腾讯网

